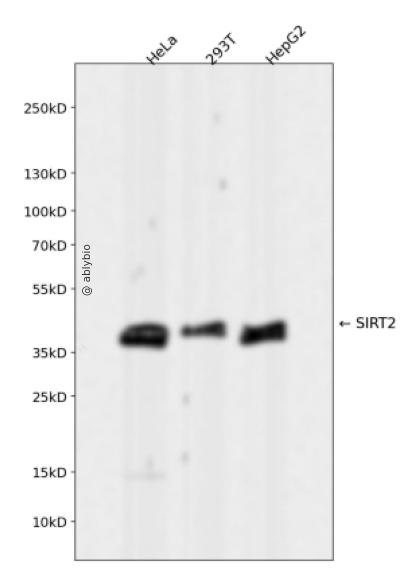
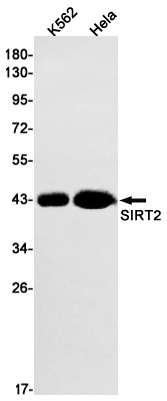
Western blot analysis of SIRT2 expressed in HeLa,293T,HepG2 using SIRT2 Rabbit mAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
Western blot analysis of SIRT2 expressed in HeLa,293T,HepG2 using SIRT2 Rabbit mAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
This gene encodes a member of the sirtuin family of proteins, homologs to the yeast Sir2 protein. Members of the sirtuin family are characterized by a sirtuin core domain and grouped into four classes. The functions of human sirtuins have not yet been determined; however, yeast sirtuin proteins are known to regulate epigenetic gene silencing and suppress recombination of rDNA. Studies suggest that the human sirtuins may function as intracellular regulatory proteins with mono-ADP-ribosyltransferase activity. The protein encoded by this gene is included in class I of the sirtuin family. Several transcript variants are resulted from alternative splicing of this gene. [provided by RefSeq, Jul 2010]
SIRT2,SIRT2 Rabbit mAb,NAD-dependent protein deacylase sirtuin-2,NAD-dependent protein defatty-acylase sirtuin-2,Regulatory protein SIR2 homolog 2,SIR2-like protein 2,SIR2L,SIR2L2
组织表达
Isoform 1 is expressed in heart, liver and skeletal muscle, weakly expressed in the cortex. Isoform 2 is strongly expressed in the cortex, weakly expressed in heart and liver. Weakly expressed in several malignancies including breast, liver, brain, kidney and prostate cancers compared to normal tissues. Weakly expressed in glioma cell lines compared to normal brain tissues (at protein level). Widely expressed. Highly expressed in heart, brain and skeletal muscle, while it is weakly expressed in placenta and lung. Down-regulated in many gliomas suggesting that it may act as a tumor suppressor gene in human gliomas possibly through the regulation of microtubule network.
功能
NAD-dependent protein deacetylase, which deacetylates internal lysines on histone and alpha-tubulin as well as many other proteins such as key transcription factors (PubMed:24177535, PubMed:12620231, PubMed:16648462, PubMed:18249187, PubMed:18332217, PubMed:18995842, PubMed:20587414, PubMed:21081649, PubMed:20543840, PubMed:22014574, PubMed:21726808, PubMed:21949390, PubMed:22771473, PubMed:23468428, PubMed:23908241, PubMed:24940000, PubMed:24769394, PubMed:24681946). Participates in the modulation of multiple and diverse biological processes such as cell cycle control, genomic integrity, microtubule dynamics, cell differentiation, metabolic networks, and autophagy. Plays a major role in the control of cell cycle progression and genomic stability. Functions in the antephase checkpoint preventing precocious mitotic entry in response to microtubule stress agents, and hence allowing proper inheritance of chromosomes. Positively regulates the anaphase promoting complex/cyclosome (APC/C) ubiquitin ligase complex activity by deacetylating CDC20 and FZR1, then allowing progression through mitosis. Associates both with chromatin at transcriptional start sites (TSSs) and enhancers of active genes. Plays a role in cell cycle and chromatin compaction through epigenetic modulation of the regulation of histone H4 'Lys-20' methylation (H4K20me1) during early mitosis. Specifically deacetylates histone H4 at 'Lys-16' (H4K16ac) between the G2/M transition and metaphase enabling H4K20me1 deposition by KMT5A leading to ulterior levels of H4K20me2 and H4K20me3 deposition throughout cell cycle, and mitotic S-phase progression (PubMed:23468428). Deacetylates KMT5A modulating KMT5A chromatin localization during the mitotic stress response (PubMed:23468428). Deacetylates also histone H3 at 'Lys-57' (H3K56ac) during the mitotic G2/M transition. Upon bacterium Listeria monocytogenes infection, deacetylates 'Lys-18' of histone H3 in a receptor tyrosine kinase MET- and PI3K/Akt-dependent manner, thereby inhibiting transcriptional activity and promoting late stages of listeria infection (PubMed:23908241). During oocyte meiosis progression, may deacetylate histone H4 at 'Lys-16' (H4K16ac) and alpha-tubulin, regulating spindle assembly and chromosome alignment by influencing microtubule dynamics and kinetochore function. Deacetylates histone H4 at 'Lys-16' (H4K16ac) at the VEGFA promoter and thereby contributes to regulate expression of VEGFA, a key regulator of angiogenesis (PubMed:24940000). Deacetylates alpha-tubulin at 'Lys-40' and hence controls neuronal motility, oligodendroglial cell arbor projection processes and proliferation of non-neuronal cells. Phosphorylation at Ser-368 by a G1/S-specific cyclin E-CDK2 complex inactivates SIRT2-mediated alpha-tubulin deacetylation, negatively regulating cell adhesion, cell migration and neurite outgrowth during neuronal differentiation. Deacetylates PARD3 and participates in the regulation of Schwann cell peripheral myelination formation during early postnatal development and during postinjury remyelination. Involved in several cellular metabolic pathways. Plays a role in the regulation of blood glucose homeostasis by deacetylating and stabilizing phosphoenolpyruvate carboxykinase PCK1 activity in response to low nutrient availability. Acts as a key regulator in the pentose phosphate pathway (PPP) by deacetylating and activating the glucose-6-phosphate G6PD enzyme, and therefore, stimulates the production of cytosolic NADPH to counteract oxidative damage. Maintains energy homeostasis in response to nutrient deprivation as well as energy expenditure by inhibiting adipogenesis and promoting lipolysis. Attenuates adipocyte differentiation by deacetylating and promoting FOXO1 interaction to PPARG and subsequent repression of PPARG-dependent transcriptional activity. Plays a role in the regulation of lysosome-mediated degradation of protein aggregates by autophagy in neuronal cells. Deacetylates FOXO1 in response to oxidative stress or serum deprivation, thereby negatively regulating FOXO1-mediated autophagy (PubMed:20543840). Deacetylates a broad range of transcription factors and co-regulators regulating target gene expression. Deacetylates transcriptional factor FOXO3 stimulating the ubiquitin ligase SCF(SKP2)-mediated FOXO3 ubiquitination and degradation (By similarity). Deacetylates HIF1A and therefore promotes HIF1A degradation and inhibition of HIF1A transcriptional activity in tumor cells in response to hypoxia (PubMed:24681946). Deacetylates RELA in the cytoplasm inhibiting NF-kappaB-dependent transcription activation upon TNF-alpha stimulation. Inhibits transcriptional activation by deacetylating p53/TP53 and EP300 (PubMed:18249187). Deacetylates also EIF5A (PubMed:22771473). Functions as a negative regulator on oxidative stress-tolerance in response to anoxia-reoxygenation conditions. Plays a role as tumor suppressor (PubMed:22014574).
a. 贴壁培养细胞收集
去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
b. 悬浮培养细胞收集
速离心悬浮细胞,弃上清,收集沉淀。手指轻弹细胞,使其松散。
c. 组织样本收集
把组织剪切成细小的碎片,越小越好。取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
(2)总蛋白提取
a. 细胞/组织裂解
将装有细胞沉淀或组织碎片的容器完全插入冰中。细胞沉淀按照1mL裂解液/107个细胞(1个T75培养瓶细胞量)的比例加入相应体积的裂解液(细胞量足够时都加入3mL,不足时根据细胞量计算),裂解20min,每隔5min将离心管置于涡旋振荡仪上震荡10s。组织碎片按照0.5mL 裂解液/100mg组织向匀浆器中加入蛋白裂解液,每3min研磨一次,重复5次,使组织尽量碾碎。(裂解液中根据需要选择添加或不添加蛋白酶抑制剂)。
b. 离心
把裂解好的样品配平后,置于预冷的高速冷冻离心机中,12000 rpm,15min。
c. 蛋白变性
完成离心后,上清即为蛋白提取液。吸取少量蛋白提取液做蛋白浓度测定。向剩余的蛋白提取液的离心管中加入1/5上清体积的5×Loading Buffer(最终工作液为1X),待干式恒温器温度升至95℃后,将1.5mL离心管插入加热孔中,95℃加热变性10min,待液体完全冷却后置于-20℃保存。
(3)蛋白浓度测定(BCA法)
a. BCA工作液的配置
根据样品数量,按50体积BCA试剂A加入1体积BCA试剂B(50:1)配置适量BCA工作液,充分混匀。BCA工作液室温24h内稳定。
b. 标准品测定
取10μl蛋白标准品(5mg/ml BSA)稀释至50μl,使终浓度为1mg/ml。稀释后的蛋白标准品可以-20℃长期保存。此标准品溶液的稀释液可使用去离子水或1*PBS。将标准品按0、1、2、4、8、12、16、20μl加入到96孔板中,加稀释液补足到20μl(见附表)。加适当体积样品到96孔板的样品孔中,如果样本不足20μl,需加稀释液补足到20μl。请注意记录样品体积。各孔加入200μl BCA工作液,37℃放置20-30min。用酶标仪测定A562,或540-595nm之间的其他波长吸光度。根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
a. Western Transfer Buffer至少提前2h (即开始电泳后)放入-20℃冰箱预冷,但注意避免结冰。
b. 根据胶体大小,将Filter Paper及Nitrocellulose membrane剪裁至合适尺寸。
c. 目的蛋白>20KD选择0.45μm NC膜/PVDF膜;目的蛋白<20KD选择0.2μmNC膜或PVDF膜,选择完毕后将NC膜放在Western Transfer Buffer中浸泡备用,注意如使用的是PVDF膜需先放入甲醇中浸泡5-10min,再放入Western Transfer Buffer中浸泡备用。
(2)裂解液&洗杂液:Cell lysis buffer for IP (without inhibitors)
(3)蛋白酶抑制剂
(4)封闭液:含 3% BSA 的 1X PBS
(5)1×PBS 缓冲液
(6)5×loding buffer(使用时用去离子水稀释至工作浓度即可)
(7)Control IgG (AC005/ AC011/AC034)
二、实验步骤
1、样本处理
(1)贴壁培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
c. 按照6孔板每孔加入100~200μl裂解液的比例,加入裂解液。移液器轻轻吹打,使裂解液和细胞充分接触。通常裂解液作用于细胞1~5s内,细胞会被裂解。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(2)悬浮培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 速离心悬浮细胞,弃上清,收集沉淀。
c. 手指轻弹细胞,使其松散。按照6孔板每孔加入100~200μl裂解液的比例,加入NP-40裂解液。通常6孔板每孔加入100~200μl裂解液已经足够,但如果细胞密度非常高可以适当加大裂解液的用量150~200μl,再用手指轻弹以充分裂解细胞。充分裂解后应无明显沉淀。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(3)组织样本
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 把组织剪切成细小的碎片,越小越好。
c. 取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
d. 按照每20mg组织加入100~200μl裂解液的比例,加入含有PMSF的裂解液。冰上或4℃裂解30-60min。(步骤3、4也可采用以下过程:按照每20mg组织加入100~200μl裂解液的比例加入NP-40裂解液。用玻璃匀浆器或组织研磨器匀浆,直至充分裂解,过程尽量控制在1~2min之内,以减少蛋白的降解。)
e. 按照每20mg组织加入100~200μl裂解液的比例,加入裂解液。
f. 1000~12000g,4℃离心10~15min(如无低温离心机,室温下离心也可),取上清。
2、磁珠预处理
(1)将rProtein A/G Plus MaqPoly Beads颠倒或漩涡混匀,翻转瓶身发现底部无黑色沉淀即可。
(2)取30μl rProtein A/G Plus MaqPoly Beads至新的EP管中,放在磁分离器上,待溶液澄清后,用移液器吸弃保护液。
(3)将EP管从磁分离器上取下来,加入1ml Cell lysis buffer for IP (without inhibitors),混匀,放置在磁分离器上,收集磁珠,用移液器吸弃洗杂液,重复2次。