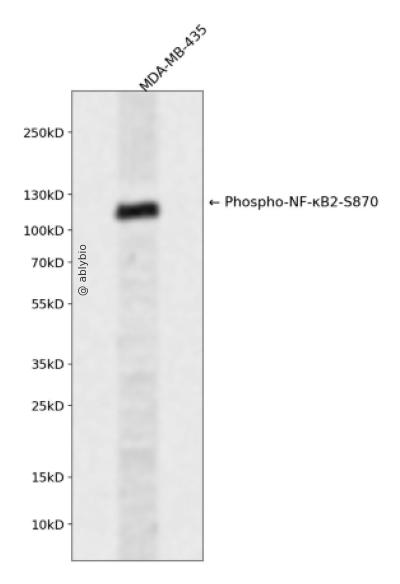
Western blot analysis of Phospho-NF-κB2-S870 expressed in MDA-MB-435 using Phospho-NF-κB2-S870 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
Western blot analysis of Phospho-NF-κB2-S870 expressed in MDA-MB-435 using Phospho-NF-κB2-S870 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
This gene encodes a subunit of the transcription factor complex nuclear factor-kappa-B (NFkB). The NFkB complex is expressed in numerous cell types and functions as a central activator of genes involved in inflammation and immune function. The protein encoded by this gene can function as both a transcriptional activator or repressor depending on its dimerization partner. The p100 full-length protein is co-translationally processed into a p52 active form. Chromosomal rearrangements and translocations of this locus have been observed in B cell lymphomas, some of which may result in the formation of fusion proteins. There is a pseudogene for this gene on chromosome 18. Alternative splicing results in multiple transcript variants.
NF-kappa-B is a pleiotropic transcription factor present in almost all cell types and is the endpoint of a series of signal transduction events that are initiated by a vast array of stimuli related to many biological processes such as inflammation, immunity, differentiation, cell growth, tumorigenesis and apoptosis. NF-kappa-B is a homo- or heterodimeric complex formed by the Rel-like domain-containing proteins RELA/p65, RELB, NFKB1/p105, NFKB1/p50, REL and NFKB2/p52. The dimers bind at kappa-B sites in the DNA of their target genes and the individual dimers have distinct preferences for different kappa-B sites that they can bind with distinguishable affinity and specificity. Different dimer combinations act as transcriptional activators or repressors, respectively. NF-kappa-B is controlled by various mechanisms of post-translational modification and subcellular compartmentalization as well as by interactions with other cofactors or corepressors. NF-kappa-B complexes are held in the cytoplasm in an inactive state complexed with members of the NF-kappa-B inhibitor (I-kappa-B) family. In a conventional activation pathway, I-kappa-B is phosphorylated by I-kappa-B kinases (IKKs) in response to different activators, subsequently degraded thus liberating the active NF-kappa-B complex which translocates to the nucleus. In a non-canonical activation pathway, the MAP3K14-activated CHUK/IKKA homodimer phosphorylates NFKB2/p100 associated with RelB, inducing its proteolytic processing to NFKB2/p52 and the formation of NF-kappa-B RelB-p52 complexes. The NF-kappa-B heterodimeric RelB-p52 complex is a transcriptional activator. The NF-kappa-B p52-p52 homodimer is a transcriptional repressor. NFKB2 appears to have dual functions such as cytoplasmic retention of attached NF-kappa-B proteins by p100 and generation of p52 by a cotranslational processing. The proteasome-mediated process ensures the production of both p52 and p100 and preserves their independent function. p52 binds to the kappa-B consensus sequence 5'-GGRNNYYCC-3', located in the enhancer region of genes involved in immune response and acute phase reactions. p52 and p100 are respectively the minor and major form; the processing of p100 being relatively poor. Isoform p49 is a subunit of the NF-kappa-B protein complex, which stimulates the HIV enhancer in synergy with p65. In concert with RELB, regulates the circadian clock by repressing the transcriptional activator activity of the CLOCK-ARNTL/BMAL1 heterodimer.
a. 贴壁培养细胞收集
去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
b. 悬浮培养细胞收集
速离心悬浮细胞,弃上清,收集沉淀。手指轻弹细胞,使其松散。
c. 组织样本收集
把组织剪切成细小的碎片,越小越好。取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
(2)总蛋白提取
a. 细胞/组织裂解
将装有细胞沉淀或组织碎片的容器完全插入冰中。细胞沉淀按照1mL裂解液/107个细胞(1个T75培养瓶细胞量)的比例加入相应体积的裂解液(细胞量足够时都加入3mL,不足时根据细胞量计算),裂解20min,每隔5min将离心管置于涡旋振荡仪上震荡10s。组织碎片按照0.5mL 裂解液/100mg组织向匀浆器中加入蛋白裂解液,每3min研磨一次,重复5次,使组织尽量碾碎。(裂解液中根据需要选择添加或不添加蛋白酶抑制剂)。
b. 离心
把裂解好的样品配平后,置于预冷的高速冷冻离心机中,12000 rpm,15min。
c. 蛋白变性
完成离心后,上清即为蛋白提取液。吸取少量蛋白提取液做蛋白浓度测定。向剩余的蛋白提取液的离心管中加入1/5上清体积的5×Loading Buffer(最终工作液为1X),待干式恒温器温度升至95℃后,将1.5mL离心管插入加热孔中,95℃加热变性10min,待液体完全冷却后置于-20℃保存。
(3)蛋白浓度测定(BCA法)
a. BCA工作液的配置
根据样品数量,按50体积BCA试剂A加入1体积BCA试剂B(50:1)配置适量BCA工作液,充分混匀。BCA工作液室温24h内稳定。
b. 标准品测定
取10μl蛋白标准品(5mg/ml BSA)稀释至50μl,使终浓度为1mg/ml。稀释后的蛋白标准品可以-20℃长期保存。此标准品溶液的稀释液可使用去离子水或1*PBS。将标准品按0、1、2、4、8、12、16、20μl加入到96孔板中,加稀释液补足到20μl(见附表)。加适当体积样品到96孔板的样品孔中,如果样本不足20μl,需加稀释液补足到20μl。请注意记录样品体积。各孔加入200μl BCA工作液,37℃放置20-30min。用酶标仪测定A562,或540-595nm之间的其他波长吸光度。根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
a. Western Transfer Buffer至少提前2h (即开始电泳后)放入-20℃冰箱预冷,但注意避免结冰。
b. 根据胶体大小,将Filter Paper及Nitrocellulose membrane剪裁至合适尺寸。
c. 目的蛋白>20KD选择0.45μm NC膜/PVDF膜;目的蛋白<20KD选择0.2μmNC膜或PVDF膜,选择完毕后将NC膜放在Western Transfer Buffer中浸泡备用,注意如使用的是PVDF膜需先放入甲醇中浸泡5-10min,再放入Western Transfer Buffer中浸泡备用。
(2)裂解液&洗杂液:Cell lysis buffer for IP (without inhibitors)
(3)蛋白酶抑制剂
(4)封闭液:含 3% BSA 的 1X PBS
(5)1×PBS 缓冲液
(6)5×loding buffer(使用时用去离子水稀释至工作浓度即可)
(7)Control IgG (AC005/ AC011/AC034)
二、实验步骤
1、样本处理
(1)贴壁培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
c. 按照6孔板每孔加入100~200μl裂解液的比例,加入裂解液。移液器轻轻吹打,使裂解液和细胞充分接触。通常裂解液作用于细胞1~5s内,细胞会被裂解。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(2)悬浮培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 速离心悬浮细胞,弃上清,收集沉淀。
c. 手指轻弹细胞,使其松散。按照6孔板每孔加入100~200μl裂解液的比例,加入NP-40裂解液。通常6孔板每孔加入100~200μl裂解液已经足够,但如果细胞密度非常高可以适当加大裂解液的用量150~200μl,再用手指轻弹以充分裂解细胞。充分裂解后应无明显沉淀。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(3)组织样本
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 把组织剪切成细小的碎片,越小越好。
c. 取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
d. 按照每20mg组织加入100~200μl裂解液的比例,加入含有PMSF的裂解液。冰上或4℃裂解30-60min。(步骤3、4也可采用以下过程:按照每20mg组织加入100~200μl裂解液的比例加入NP-40裂解液。用玻璃匀浆器或组织研磨器匀浆,直至充分裂解,过程尽量控制在1~2min之内,以减少蛋白的降解。)
e. 按照每20mg组织加入100~200μl裂解液的比例,加入裂解液。
f. 1000~12000g,4℃离心10~15min(如无低温离心机,室温下离心也可),取上清。
2、磁珠预处理
(1)将rProtein A/G Plus MaqPoly Beads颠倒或漩涡混匀,翻转瓶身发现底部无黑色沉淀即可。
(2)取30μl rProtein A/G Plus MaqPoly Beads至新的EP管中,放在磁分离器上,待溶液澄清后,用移液器吸弃保护液。
(3)将EP管从磁分离器上取下来,加入1ml Cell lysis buffer for IP (without inhibitors),混匀,放置在磁分离器上,收集磁珠,用移液器吸弃洗杂液,重复2次。