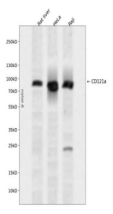
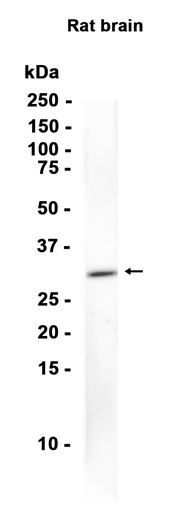
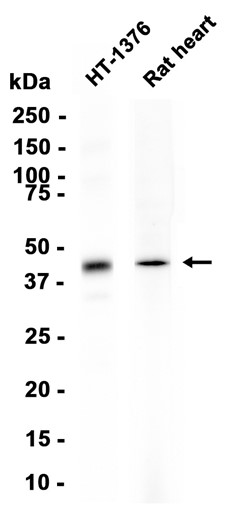
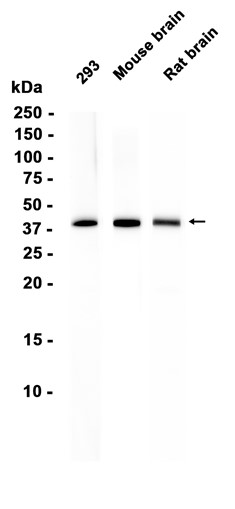
This gene is a member of the Period family of genes and is expressed in a circadian pattern in the suprachiasmatic nucleus, the primary circadian pacemaker in the mammalian brain. Genes in this family encode components of the circadian rhythms of locomotor activity, metabolism, and behavior. This gene is upregulated by CLOCK/ARNTL heterodimers but then represses this upregulation in a feedback loop using PER/CRY heterodimers to interact with CLOCK/ARNTL. Polymorphisms in this gene may increase the risk of getting certain cancers and have been linked to sleep disorders.
PER2,FASPS,FASPS1,PER2 (Phospho-Ser662) Antibody,Circadian clock protein PERIOD 2,KIAA0347,PER2 (Phospho-Ser662)
组织表达
Widely expressed. Found in heart, brain, placenta, lung, liver, skeleatal muscle, kidney and pancreas. High levels in skeletal muscle and pancreas. Low levels in lung. Isoform 2 is expressed in keratinocytes (at protein level).
功能
Transcriptional repressor which forms a core component of the circadian clock. The circadian clock, an internal time-keeping system, regulates various physiological processes through the generation of approximately 24 hour circadian rhythms in gene expression, which are translated into rhythms in metabolism and behavior. It is derived from the Latin roots 'circa' (about) and 'diem' (day) and acts as an important regulator of a wide array of physiological functions including metabolism, sleep, body temperature, blood pressure, endocrine, immune, cardiovascular, and renal function. Consists of two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system. Both the central and peripheral clocks can be reset by environmental cues, also known as Zeitgebers (German for 'timegivers'). The predominant Zeitgeber for the central clock is light, which is sensed by retina and signals directly to the SCN. The central clock entrains the peripheral clocks through neuronal and hormonal signals, body temperature and feeding-related cues, aligning all clocks with the external light/dark cycle. Circadian rhythms allow an organism to achieve temporal homeostasis with its environment at the molecular level by regulating gene expression to create a peak of protein expression once every 24 hours to control when a particular physiological process is most active with respect to the solar day. Transcription and translation of core clock components (CLOCK, NPAS2, ARNTL/BMAL1, ARNTL2/BMAL2, PER1, PER2, PER3, CRY1 and CRY2) plays a critical role in rhythm generation, whereas delays imposed by post-translational modifications (PTMs) are important for determining the period (tau) of the rhythms (tau refers to the period of a rhythm and is the length, in time, of one complete cycle). A diurnal rhythm is synchronized with the day/night cycle, while the ultradian and infradian rhythms have a period shorter and longer than 24 hours, respectively. Disruptions in the circadian rhythms contribute to the pathology of cardiovascular diseases, cancer, metabolic syndrome and aging. A transcription/translation feedback loop (TTFL) forms the core of the molecular circadian clock mechanism. Transcription factors, CLOCK or NPAS2 and ARNTL/BMAL1 or ARNTL2/BMAL2, form the positive limb of the feedback loop, act in the form of a heterodimer and activate the transcription of core clock genes and clock-controlled genes (involved in key metabolic processes), harboring E-box elements (5'-CACGTG-3') within their promoters. The core clock genes: PER1/2/3 and CRY1/2 which are transcriptional repressors form the negative limb of the feedback loop and interact with the CLOCK|NPAS2-ARNTL/BMAL1|ARNTL2/BMAL2 heterodimer inhibiting its activity and thereby negatively regulating their own expression. This heterodimer also activates nuclear receptors NR1D1/2 and RORA/B/G, which form a second feedback loop and which activate and repress ARNTL/BMAL1 transcription, respectively. PER1 and PER2 proteins transport CRY1 and CRY2 into the nucleus with appropriate circadian timing, but also contribute directly to repression of clock-controlled target genes through interaction with several classes of RNA-binding proteins, helicases and others transcriptional repressors. PER appears to regulate circadian control of transcription by at least three different modes. First, interacts directly with the CLOCK-ARTNL/BMAL1 at the tail end of the nascent transcript peak to recruit complexes containing the SIN3-HDAC that remodel chromatin to repress transcription. Second, brings H3K9 methyltransferases such as SUV39H1 and SUV39H2 to the E-box elements of the circadian target genes, like PER2 itself or PER1. The recruitment of each repressive modifier to the DNA seems to be very precisely temporally orchestrated by the large PER complex, the deacetylases acting before than the methyltransferases. Additionally, large PER complexes are also recruited to the target genes 3' termination site through interactions with RNA-binding proteins and helicases that may play a role in transcription termination to regulate transcription independently of CLOCK-ARTNL/BMAL1 interactions. Recruitment of large PER complexes to the elongating polymerase at PER and CRY termination sites inhibited SETX action, impeding RNA polymerase II release and thereby repressing transcriptional reinitiation. May propagate clock information to metabolic pathways via the interaction with nuclear receptors. Coactivator of PPARA and corepressor of NR1D1, binds rhythmically at the promoter of nuclear receptors target genes like ARNTL or G6PC. Directly and specifically represses PPARG proadipogenic activity by blocking PPARG recruitment to target promoters and thereby inhibiting transcriptional activation. Required for fatty acid and lipid metabolism, is involved as well in the regulation of circulating insulin levels. Plays an important role in the maintenance of cardiovascular functions through the regulation of NO and vasodilatatory prostaglandins production in aortas. Controls circadian glutamate uptake in synaptic vesicles through the regulation of VGLUT1 expression. May also be involved in the regulation of inflammatory processes. Represses the CLOCK-ARNTL/BMAL1 induced transcription of BHLHE40/DEC1 and ATF4. Negatively regulates the formation of the TIMELESS-CRY1 complex by competing with TIMELESS for binding to CRY1.
a. 贴壁培养细胞收集
去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
b. 悬浮培养细胞收集
速离心悬浮细胞,弃上清,收集沉淀。手指轻弹细胞,使其松散。
c. 组织样本收集
把组织剪切成细小的碎片,越小越好。取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
(2)总蛋白提取
a. 细胞/组织裂解
将装有细胞沉淀或组织碎片的容器完全插入冰中。细胞沉淀按照1mL裂解液/107个细胞(1个T75培养瓶细胞量)的比例加入相应体积的裂解液(细胞量足够时都加入3mL,不足时根据细胞量计算),裂解20min,每隔5min将离心管置于涡旋振荡仪上震荡10s。组织碎片按照0.5mL 裂解液/100mg组织向匀浆器中加入蛋白裂解液,每3min研磨一次,重复5次,使组织尽量碾碎。(裂解液中根据需要选择添加或不添加蛋白酶抑制剂)。
b. 离心
把裂解好的样品配平后,置于预冷的高速冷冻离心机中,12000 rpm,15min。
c. 蛋白变性
完成离心后,上清即为蛋白提取液。吸取少量蛋白提取液做蛋白浓度测定。向剩余的蛋白提取液的离心管中加入1/5上清体积的5×Loading Buffer(最终工作液为1X),待干式恒温器温度升至95℃后,将1.5mL离心管插入加热孔中,95℃加热变性10min,待液体完全冷却后置于-20℃保存。
(3)蛋白浓度测定(BCA法)
a. BCA工作液的配置
根据样品数量,按50体积BCA试剂A加入1体积BCA试剂B(50:1)配置适量BCA工作液,充分混匀。BCA工作液室温24h内稳定。
b. 标准品测定
取10μl蛋白标准品(5mg/ml BSA)稀释至50μl,使终浓度为1mg/ml。稀释后的蛋白标准品可以-20℃长期保存。此标准品溶液的稀释液可使用去离子水或1*PBS。将标准品按0、1、2、4、8、12、16、20μl加入到96孔板中,加稀释液补足到20μl(见附表)。加适当体积样品到96孔板的样品孔中,如果样本不足20μl,需加稀释液补足到20μl。请注意记录样品体积。各孔加入200μl BCA工作液,37℃放置20-30min。用酶标仪测定A562,或540-595nm之间的其他波长吸光度。根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
a. Western Transfer Buffer至少提前2h (即开始电泳后)放入-20℃冰箱预冷,但注意避免结冰。
b. 根据胶体大小,将Filter Paper及Nitrocellulose membrane剪裁至合适尺寸。
c. 目的蛋白>20KD选择0.45μm NC膜/PVDF膜;目的蛋白<20KD选择0.2μmNC膜或PVDF膜,选择完毕后将NC膜放在Western Transfer Buffer中浸泡备用,注意如使用的是PVDF膜需先放入甲醇中浸泡5-10min,再放入Western Transfer Buffer中浸泡备用。
(2)裂解液&洗杂液:Cell lysis buffer for IP (without inhibitors)
(3)蛋白酶抑制剂
(4)封闭液:含 3% BSA 的 1X PBS
(5)1×PBS 缓冲液
(6)5×loding buffer(使用时用去离子水稀释至工作浓度即可)
(7)Control IgG (AC005/ AC011/AC034)
二、实验步骤
1、样本处理
(1)贴壁培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
c. 按照6孔板每孔加入100~200μl裂解液的比例,加入裂解液。移液器轻轻吹打,使裂解液和细胞充分接触。通常裂解液作用于细胞1~5s内,细胞会被裂解。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(2)悬浮培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 速离心悬浮细胞,弃上清,收集沉淀。
c. 手指轻弹细胞,使其松散。按照6孔板每孔加入100~200μl裂解液的比例,加入NP-40裂解液。通常6孔板每孔加入100~200μl裂解液已经足够,但如果细胞密度非常高可以适当加大裂解液的用量150~200μl,再用手指轻弹以充分裂解细胞。充分裂解后应无明显沉淀。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(3)组织样本
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 把组织剪切成细小的碎片,越小越好。
c. 取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
d. 按照每20mg组织加入100~200μl裂解液的比例,加入含有PMSF的裂解液。冰上或4℃裂解30-60min。(步骤3、4也可采用以下过程:按照每20mg组织加入100~200μl裂解液的比例加入NP-40裂解液。用玻璃匀浆器或组织研磨器匀浆,直至充分裂解,过程尽量控制在1~2min之内,以减少蛋白的降解。)
e. 按照每20mg组织加入100~200μl裂解液的比例,加入裂解液。
f. 1000~12000g,4℃离心10~15min(如无低温离心机,室温下离心也可),取上清。
2、磁珠预处理
(1)将rProtein A/G Plus MaqPoly Beads颠倒或漩涡混匀,翻转瓶身发现底部无黑色沉淀即可。
(2)取30μl rProtein A/G Plus MaqPoly Beads至新的EP管中,放在磁分离器上,待溶液澄清后,用移液器吸弃保护液。
(3)将EP管从磁分离器上取下来,加入1ml Cell lysis buffer for IP (without inhibitors),混匀,放置在磁分离器上,收集磁珠,用移液器吸弃洗杂液,重复2次。