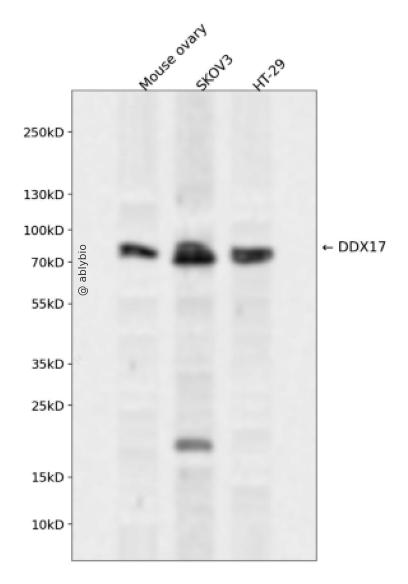
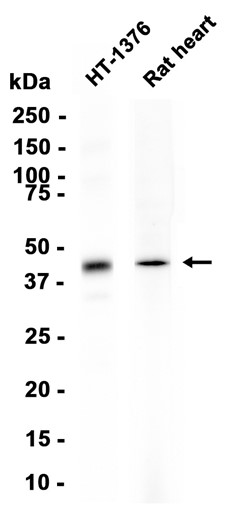
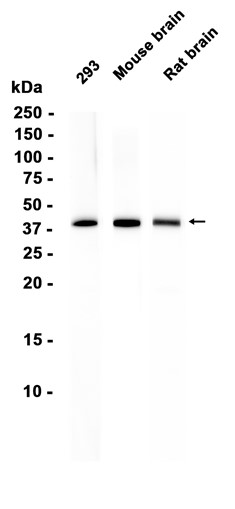
Western blot analysis of DDX17 expressed in Mouse ovary,SKOV3,HT-29 using DDX17 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
Western blot analysis of DDX17 expressed in Mouse ovary,SKOV3,HT-29 using DDX17 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
DEAD box proteins, characterized by the conserved motif Asp-Glu-Ala-Asp (DEAD), are putative RNA helicases. They are implicated in a number of cellular processes involving alteration of RNA secondary structure, such as translation initiation, nuclear and mitochondrial splicing, and ribosome and splicesosome assembly. Based on their distribution patterns, some members of this family are believed to be involved in embryogenesis, spermatogenesis, and cellular growth and division. This gene encodes a DEAD box protein, which is an ATPase activated by a variety of RNA species, but not by dsDNA. This protein, and that encoded by DDX5 gene, are more closely related to each other than to any other member of the DEAD box family. This gene can encode multiple isoforms due to both alternative splicing and the use of alternative translation initiation codons, including a non-AUG (CUG) start codon.
DDX17,P72,RH70,DDX17 Rabbit pAb,DEAD box protein 17,DEAD box protein p72,DEAD box protein p82,RNA-dependent helicase p72
组织表达
Widely expressed (PubMed:8871553). Low expression, if any, in normal colonic epithelial cells (at protein level). Levels tend to increase during colon cancer progression, from very low in benign hyperplastic polyps to very high in tubular and villous adenomas (PubMed:17699760).
功能
As an RNA helicase, unwinds RNA and alters RNA structures through ATP binding and hydrolysis. Involved in multiple cellular processes, including pre-mRNA splicing, alternative splicing, ribosomal RNA processing and miRNA processing, as well as transcription regulation. Regulates the alternative splicing of exons exhibiting specific features (PubMed:12138182, PubMed:23022728, PubMed:24910439, PubMed:22266867). For instance, promotes the inclusion of AC-rich alternative exons in CD44 transcripts (PubMed:12138182). This function requires the RNA helicase activity (PubMed:12138182, PubMed:23022728, PubMed:24910439, PubMed:22266867). Affects NFAT5 and histone macro-H2A.1/H2AFY alternative splicing in a CDK9-dependent manner (PubMed:26209609, PubMed:22266867). In NFAT5, promotes the introduction of alternative exon 4, which contains 2 stop codons and may target NFAT5 exon 4-containing transcripts to nonsense-mediated mRNA decay, leading to the down-regulation of NFAT5 protein (PubMed:22266867). Affects splicing of mediators of steroid hormone signaling pathway, including kinases that phosphorylates ESR1, such as CDK2, MAPK1 and GSK3B, and transcriptional regulators, such as CREBBP, MED1, NCOR1 and NCOR2. By affecting GSK3B splicing, participates in ESR1 and AR stabilization (PubMed:24275493). In myoblasts and epithelial cells, cooperates with HNRNPH1 to control the splicing of specific subsets of exons (PubMed:24910439). In addition to binding mature mRNAs, also interacts with certain pri-microRNAs, including MIR663/miR-663a, MIR99B/miR-99b, and MIR6087/miR-6087 (PubMed:25126784). Binds pri-microRNAs on the 3' segment flanking the stem loop via the 5'-[ACG]CAUC[ACU]-3' consensus sequence (PubMed:24581491). Required for the production of subsets of microRNAs, including MIR21 and MIR125B1 (PubMed:24581491, PubMed:27478153). May be involved not only in microRNA primary transcript processing, but also stabilization (By similarity). Participates in MYC down-regulation at high cell density through the production of MYC-targeting microRNAs (PubMed:24581491). Along with DDX5, may be involved in the processing of the 32S intermediate into the mature 28S ribosomal RNA (PubMed:17485482). Promoter-specific transcription regulator, functioning as a coactivator or corepressor depending on the context of the promoter and the transcriptional complex in which it exists (PubMed:15298701). Enhances NFAT5 transcriptional activity (PubMed:22266867). Synergizes with TP53 in the activation of the MDM2 promoter; this activity requires acetylation on lysine residues (PubMed:17226766, PubMed:20663877, PubMed:19995069). May also coactivate MDM2 transcription through a TP53-independent pathway (PubMed:17226766). Coactivates MMP7 transcription (PubMed:17226766). Along with CTNNB1, coactivates MYC, JUN, FOSL1 and cyclin D1/CCND1 transcription (PubMed:17699760). Alone or in combination with DDX5 and/or SRA1 non-coding RNA, plays a critical role in promoting the assembly of proteins required for the formation of the transcription initiation complex and chromatin remodeling leading to coactivation of MYOD1-dependent transcription. This helicase-independent activity is required for skeletal muscle cells to properly differentiate into myotubes (PubMed:17011493, PubMed:24910439). During epithelial-to-mesenchymal transition, coregulates SMAD-dependent transcriptional activity, directly controlling key effectors of differentiation, including miRNAs which in turn directly repress its expression (PubMed:24910439). Plays a role in estrogen and testosterone signaling pathway at several levels. Mediates the use of alternative promoters in estrogen-responsive genes and regulates transcription and splicing of a large number of steroid hormone target genes (PubMed:24275493, PubMed:20406972, PubMed:20663877, PubMed:19995069). Contrary to splicing regulation activity, transcriptional coregulation of the estrogen receptor ESR1 is helicase-independent (PubMed:19718048, PubMed:24275493). Plays a role in innate immunity. Specifically restricts bunyavirus infection, including Rift Valley fever virus (RVFV) or La Crosse virus (LACV), but not vesicular stomatitis virus (VSV), in an interferon- and DROSHA-independent manner (PubMed:25126784). Binds to RVFV RNA, likely via structured viral RNA elements (PubMed:25126784). Promotes mRNA degradation mediated by the antiviral zinc-finger protein ZC3HAV1, in an ATPase-dependent manner (PubMed:18334637).
a. 贴壁培养细胞收集
去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
b. 悬浮培养细胞收集
速离心悬浮细胞,弃上清,收集沉淀。手指轻弹细胞,使其松散。
c. 组织样本收集
把组织剪切成细小的碎片,越小越好。取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
(2)总蛋白提取
a. 细胞/组织裂解
将装有细胞沉淀或组织碎片的容器完全插入冰中。细胞沉淀按照1mL裂解液/107个细胞(1个T75培养瓶细胞量)的比例加入相应体积的裂解液(细胞量足够时都加入3mL,不足时根据细胞量计算),裂解20min,每隔5min将离心管置于涡旋振荡仪上震荡10s。组织碎片按照0.5mL 裂解液/100mg组织向匀浆器中加入蛋白裂解液,每3min研磨一次,重复5次,使组织尽量碾碎。(裂解液中根据需要选择添加或不添加蛋白酶抑制剂)。
b. 离心
把裂解好的样品配平后,置于预冷的高速冷冻离心机中,12000 rpm,15min。
c. 蛋白变性
完成离心后,上清即为蛋白提取液。吸取少量蛋白提取液做蛋白浓度测定。向剩余的蛋白提取液的离心管中加入1/5上清体积的5×Loading Buffer(最终工作液为1X),待干式恒温器温度升至95℃后,将1.5mL离心管插入加热孔中,95℃加热变性10min,待液体完全冷却后置于-20℃保存。
(3)蛋白浓度测定(BCA法)
a. BCA工作液的配置
根据样品数量,按50体积BCA试剂A加入1体积BCA试剂B(50:1)配置适量BCA工作液,充分混匀。BCA工作液室温24h内稳定。
b. 标准品测定
取10μl蛋白标准品(5mg/ml BSA)稀释至50μl,使终浓度为1mg/ml。稀释后的蛋白标准品可以-20℃长期保存。此标准品溶液的稀释液可使用去离子水或1*PBS。将标准品按0、1、2、4、8、12、16、20μl加入到96孔板中,加稀释液补足到20μl(见附表)。加适当体积样品到96孔板的样品孔中,如果样本不足20μl,需加稀释液补足到20μl。请注意记录样品体积。各孔加入200μl BCA工作液,37℃放置20-30min。用酶标仪测定A562,或540-595nm之间的其他波长吸光度。根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
a. Western Transfer Buffer至少提前2h (即开始电泳后)放入-20℃冰箱预冷,但注意避免结冰。
b. 根据胶体大小,将Filter Paper及Nitrocellulose membrane剪裁至合适尺寸。
c. 目的蛋白>20KD选择0.45μm NC膜/PVDF膜;目的蛋白<20KD选择0.2μmNC膜或PVDF膜,选择完毕后将NC膜放在Western Transfer Buffer中浸泡备用,注意如使用的是PVDF膜需先放入甲醇中浸泡5-10min,再放入Western Transfer Buffer中浸泡备用。
(2)裂解液&洗杂液:Cell lysis buffer for IP (without inhibitors)
(3)蛋白酶抑制剂
(4)封闭液:含 3% BSA 的 1X PBS
(5)1×PBS 缓冲液
(6)5×loding buffer(使用时用去离子水稀释至工作浓度即可)
(7)Control IgG (AC005/ AC011/AC034)
二、实验步骤
1、样本处理
(1)贴壁培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
c. 按照6孔板每孔加入100~200μl裂解液的比例,加入裂解液。移液器轻轻吹打,使裂解液和细胞充分接触。通常裂解液作用于细胞1~5s内,细胞会被裂解。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(2)悬浮培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 速离心悬浮细胞,弃上清,收集沉淀。
c. 手指轻弹细胞,使其松散。按照6孔板每孔加入100~200μl裂解液的比例,加入NP-40裂解液。通常6孔板每孔加入100~200μl裂解液已经足够,但如果细胞密度非常高可以适当加大裂解液的用量150~200μl,再用手指轻弹以充分裂解细胞。充分裂解后应无明显沉淀。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(3)组织样本
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 把组织剪切成细小的碎片,越小越好。
c. 取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
d. 按照每20mg组织加入100~200μl裂解液的比例,加入含有PMSF的裂解液。冰上或4℃裂解30-60min。(步骤3、4也可采用以下过程:按照每20mg组织加入100~200μl裂解液的比例加入NP-40裂解液。用玻璃匀浆器或组织研磨器匀浆,直至充分裂解,过程尽量控制在1~2min之内,以减少蛋白的降解。)
e. 按照每20mg组织加入100~200μl裂解液的比例,加入裂解液。
f. 1000~12000g,4℃离心10~15min(如无低温离心机,室温下离心也可),取上清。
2、磁珠预处理
(1)将rProtein A/G Plus MaqPoly Beads颠倒或漩涡混匀,翻转瓶身发现底部无黑色沉淀即可。
(2)取30μl rProtein A/G Plus MaqPoly Beads至新的EP管中,放在磁分离器上,待溶液澄清后,用移液器吸弃保护液。
(3)将EP管从磁分离器上取下来,加入1ml Cell lysis buffer for IP (without inhibitors),混匀,放置在磁分离器上,收集磁珠,用移液器吸弃洗杂液,重复2次。