Beta Arrestin 2 (YD16141) Rabbit mAb (货号:AYD11215)
A
小A
销售顾问

请使用微信扫一扫
添加顾问进行咨询
货号
AYD11215
靶点/基因
ARRB2/Arrb2
宿主
Rabbit
克隆性
Monoclonal
反应种属
Human, Mouse, Rat
应用
WB, FC, IP
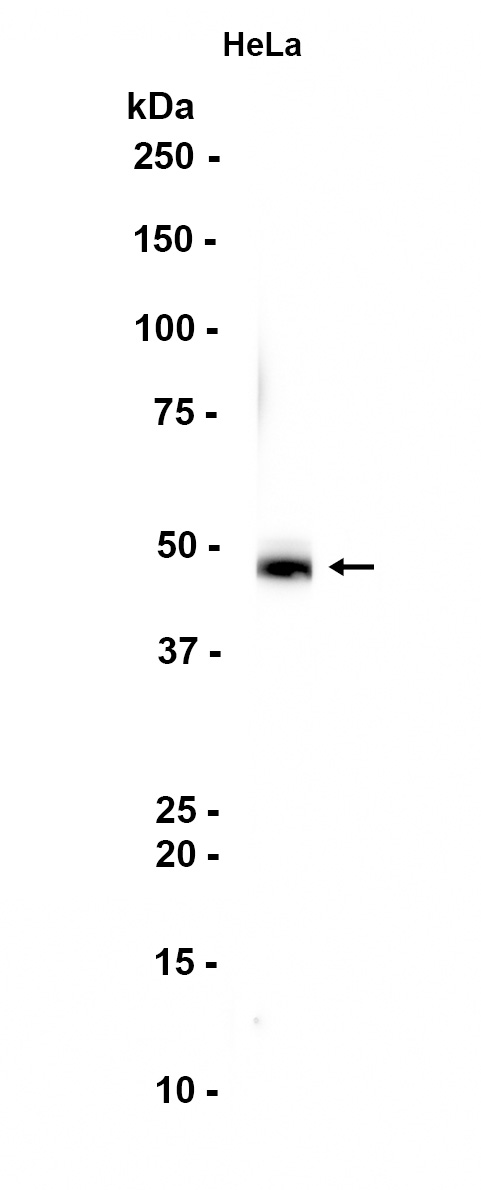
产品信息
应用指南
相关产品
抗原信息
靶点信息
资料与支持
实验步骤
常见问题
| 反应 | Human,Mouse,Rat |
|---|---|
| 宿主 | Rabbit |
| 克隆性 | Monoclonal |
| 同种型 | IgG |
| 应用 | WBFCIP |
| 推荐浓度 | |
| 理论分子量 | 46kDa/46kDa/46kDa |
| 实测分子量 | |
| 形式 | Liquid |
| 保存条件 | Store at -20℃. Avoid freeze / thaw cycles. Buffer: PBS with 0.75% BSA,50% glycerol,pH7.3. |
| 偶联物 | Unconjugated |
| 阳性对照 | |
| 细胞定位 | Cytoplasm, Nucleus, Cell membrane, Membrane, clathrin-coated pit, Cytoplasmic vesicle |
| 纯化 | 亲和纯化 |
应用与推荐条件
快速判断怎么用
以下条件基于推荐浓度、验证图说明与通用实验要求整理,可作为预实验起点;不同样本和检测体系建议做梯度优化。
WB
WB 推荐条件
| 推荐稀释 | 请参考验证图说明或咨询技术支持 |
|---|---|
| 建议样本/阳性对照 | 建议选择靶点高表达样本作为阳性对照 |
| 关键条件 | 建议使用新鲜裂解样本,按推荐稀释比例孵育一抗,并关注理论/实测分子量 |
| 预期结果 | 预期信号/条带约 46kDa/46kDa/46kDa |
| 对照设置 | 建议设置阳性样本、阴性样本和二抗/同型对照 |
FC
FC 推荐条件
| 推荐稀释 | 请参考验证图说明或咨询技术支持 |
|---|---|
| 建议样本/阳性对照 | 建议选择靶点高表达样本作为阳性对照 |
| 关键条件 | 如检测胞内靶点,需优化固定/通透条件,并设置同型对照 |
| 预期结果 | 预期阳性群体荧光信号相对阴性/同型对照右移 |
| 对照设置 | 建议设置阳性样本、阴性样本和二抗/同型对照 |
IP
IP 推荐条件
| 推荐稀释 | 请参考验证图说明或咨询技术支持 |
|---|---|
| 建议样本/阳性对照 | 建议选择靶点高表达样本作为阳性对照 |
| 关键条件 | 建议从页面推荐浓度开始,结合样本与检测体系做梯度优化 |
| 预期结果 | 预期信号/条带约 46kDa/46kDa/46kDa |
| 对照设置 | 建议设置阳性样本、阴性样本和二抗/同型对照 |
相关产品
查找相关产品 >>
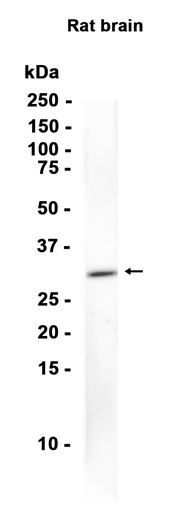
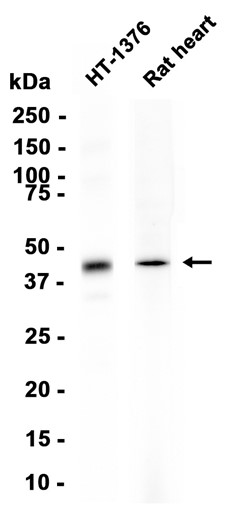
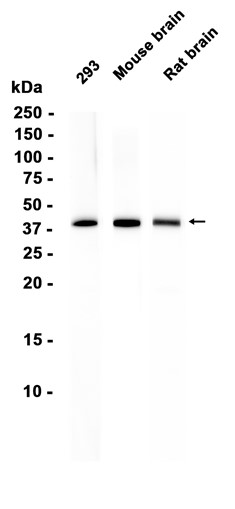
抗原信息
| 抗原信息 | 请咨询 技术支持 |
|---|---|
| 序列 | Email For Sequence |
靶点信息
| 研究背景 | Functions in regulating agonist-mediated G-protein coupled receptor (GPCR) signaling by mediating both receptor desensitization and resensitization processes (PubMed:37209686, PubMed:38175886). During homologous desensitization, beta-arrestins bind to the GPRK-phosphorylated receptor and sterically preclude its coupling to the cognate G-protein; the binding appears to require additional receptor determinants exposed only in the active receptor conformation. The beta-arrestins target many receptors for internalization by acting as endocytic adapters (CLASPs, clathrin-associated sorting proteins) and recruiting the GPRCs to the adapter protein 2 complex 2 (AP-2) in clathrin-coated pits (CCPs). However, the extent of beta-arrestin involvement appears to vary significantly depending on the receptor, agonist and cell type. Internalized arrestin-receptor complexes traffic to intracellular endosomes, where they remain uncoupled from G-proteins. Two different modes of arrestin-mediated internalization occur. Class A receptors, like ADRB2, OPRM1, ENDRA, D1AR and ADRA1B dissociate from beta-arrestin at or near the plasma membrane and undergo rapid recycling. Class B receptors, like AVPR2, AGTR1, NTSR1, TRHR and TACR1 internalize as a complex with arrestin and traffic with it to endosomal vesicles, presumably as desensitized receptors, for extended periods of time. Receptor resensitization then requires that receptor-bound arrestin is removed so that the receptor can be dephosphorylated and returned to the plasma membrane. Mediates endocytosis of CCR7 following ligation of CCL19 but not CCL21. Involved in internalization of P2RY1, P2RY4, P2RY6 and P2RY11 and ATP-stimulated internalization of P2RY2. Involved in phosphorylation-dependent internalization of OPRD1 and subsequent recycling or degradation. Involved in ubiquitination of IGF1R. Beta-arrestins function as multivalent adapter proteins that can switch the GPCR from a G-protein signaling mode that transmits short-lived signals from the plasma membrane via small molecule second messengers and ion channels to a beta-arrestin signaling mode that transmits a distinct set of signals that are initiated as the receptor internalizes and transits the intracellular compartment. Acts as a signaling scaffold for MAPK pathways such as MAPK1/3 (ERK1/2) and MAPK10 (JNK3). ERK1/2 and JNK3 activated by the beta-arrestin scaffold are largely excluded from the nucleus and confined to cytoplasmic locations such as endocytic vesicles, also called beta-arrestin signalosomes. Acts as a signaling scaffold for the AKT1 pathway. GPCRs for which the beta-arrestin-mediated signaling relies on both ARRB1 and ARRB2 (codependent regulation) include ADRB2, F2RL1 and PTH1R. For some GPCRs the beta-arrestin-mediated signaling relies on either ARRB1 or ARRB2 and is inhibited by the other respective beta-arrestin form (reciprocal regulation). Increases ERK1/2 signaling in AGTR1- and AVPR2-mediated activation (reciprocal regulation). Involved in CCR7-mediated ERK1/2 signaling involving ligand CCL19. Is involved in type-1A angiotensin II receptor/AGTR1-mediated ERK activity. Is involved in type-1A angiotensin II receptor/AGTR1-mediated MAPK10 activity. Is involved in dopamine-stimulated AKT1 activity in the striatum by disrupting the association of AKT1 with its negative regulator PP2A. Involved in AGTR1-mediated chemotaxis. Appears to function as signaling scaffold involved in regulation of MIP-1-beta-stimulated CCR5-dependent chemotaxis. Involved in attenuation of NF-kappa-B-dependent transcription in response to GPCR or cytokine stimulation by interacting with and stabilizing CHUK. Suppresses UV-induced NF-kappa-B-dependent activation by interacting with CHUK. The function is promoted by stimulation of ADRB2 and dephosphorylation of ARRB2. Involved in p53/TP53-mediated apoptosis by regulating MDM2 and reducing the MDM2-mediated degradation of p53/TP53. May serve as nuclear messenger for GPCRs. Upon stimulation of OR1D2, may be involved in regulation of gene expression during the early processes of fertilization. Also involved in regulation of receptors other than GPCRs. Involved in endocytosis of TGFBR2 and TGFBR3 and down-regulates TGF-beta signaling such as NF-kappa-B activation. Involved in endocytosis of low-density lipoprotein receptor/LDLR. Involved in endocytosis of smoothened homolog/Smo, which also requires GRK2. Involved in endocytosis of SLC9A5. Involved in endocytosis of ENG and subsequent TGF-beta-mediated ERK activation and migration of epithelial cells. Involved in Toll-like receptor and IL-1 receptor signaling through the interaction with TRAF6 which prevents TRAF6 autoubiquitination and oligomerization required for activation of NF-kappa-B and JUN (PubMed:26839314). Involved in insulin resistance by acting as insulin-induced signaling scaffold for SRC, AKT1 and INSR. Involved in regulation of inhibitory signaling of natural killer cells by recruiting PTPN6 and PTPN11 to KIR2DL1. Involved in IL8-mediated granule release in neutrophils. Involved in the internalization of the atypical chemokine receptor ACKR3. Acts as an adapter protein coupling FFAR4 receptor to specific downstream signaling pathways, as well as mediating receptor endocytosis (PubMed:22282525, PubMed:23809162). During the activation step of NLRP3 inflammasome, directly associates with NLRP3 leading to inhibition of pro-inflammatory cytokine release and inhibition of inflammation (PubMed:23809162) Functions in regulating agonist-mediated G-protein coupled receptor (GPCR) signaling by mediating both receptor desensitization and resensitization processes. During homologous desensitization, beta-arrestins bind to the GPRK-phosphorylated receptor and sterically preclude its coupling to the cognate G-protein; the binding appears to require additional receptor determinants exposed only in the active receptor conformation. The beta-arrestins target many receptors for internalization by acting as endocytic adapters (CLASPs, clathrin-associated sorting proteins) and recruiting the GPRCs to the adapter protein 2 complex 2 (AP-2) in clathrin-coated pits (CCPs). However, the extent of beta-arrestin involvement appears to vary significantly depending on the receptor, agonist and cell type. Internalized arrestin-receptor complexes traffic to intracellular endosomes, where they remain uncoupled from G-proteins. Two different modes of arrestin-mediated internalization occur. Class A receptors, like ADRB2, OPRM1, ENDRA, D1AR and ADRA1B dissociate from beta-arrestin at or near the plasma membrane and undergo rapid recycling. Class B receptors, like AVPR2, AGTR1, NTSR1, TRHR and TACR1 internalize as a complex with arrestin and traffic with it to endosomal vesicles, presumably as desensitized receptors, for extended periods of time. Receptor resensitization then requires that receptor-bound arrestin is removed so that the receptor can be dephosphorylated and returned to the plasma membrane. Mediates endocytosis of CCR7 following ligation of CCL19 but not CCL21. Involved in internalization of P2RY1, P2RY4, P2RY6 and P2RY11 and ATP-stimulated internalization of P2RY2. Involved in phosphorylation-dependent internalization of OPRD1 and subsequent recycling or degradation. Involved in ubiquitination of IGF1R. Beta-arrestins function as multivalent adapter proteins that can switch the GPCR from a G-protein signaling mode that transmits short-lived signals from the plasma membrane via small molecule second messengers and ion channels to a beta-arrestin signaling mode that transmits a distinct set of signals that are initiated as the receptor internalizes and transits the intracellular compartment. Acts as a signaling scaffold for MAPK pathways such as MAPK1/3 (ERK1/2) and MAPK10 (JNK3). ERK1/2 and JNK3 activated by the beta-arrestin scaffold are largely excluded from the nucleus and confined to cytoplasmic locations such as endocytic vesicles, also called beta-arrestin signalosomes. Acts as a signaling scaffold for the AKT1 pathway. GPCRs for which the beta-arrestin-mediated signaling relies on both ARRB1 and ARRB2 (codependent regulation) include ADRB2, F2RL1 and PTH1R. For some GPCRs the beta-arrestin-mediated signaling relies on either ARRB1 or ARRB2 and is inhibited by the other respective beta-arrestin form (reciprocal regulation). Increases ERK1/2 signaling in AGTR1- and AVPR2-mediated activation (reciprocal regulation). Involved in CCR7-mediated ERK1/2 signaling involving ligand CCL19. Is involved in type-1A angiotensin II receptor/AGTR1-mediated ERK activity. Is involved in type-1A angiotensin II receptor/AGTR1-mediated MAPK10 activity. Is involved in dopamine-stimulated AKT1 activity in the striatum by disrupting the association of AKT1 with its negative regulator PP2A. Involved in AGTR1-mediated chemotaxis. Appears to function as signaling scaffold involved in regulation of MIP-1-beta-stimulated CCR5-dependent chemotaxis. Involved in attenuation of NF-kappa-B-dependent transcription in response to GPCR or cytokine stimulation by interacting with and stabilizing CHUK. Suppresses UV-induced NF-kappa-B-dependent activation by interacting with CHUK. The function is promoted by stimulation of ADRB2 and dephosphorylation of ARRB2. Involved in IL8-mediated granule release in neutrophils (By similarity). Involved in p53/TP53-mediated apoptosis by regulating MDM2 and reducing the MDM2-mediated degradation of p53/TP53. May serve as nuclear messenger for GPCRs. Upon stimulation of OR1D2, may be involved in regulation of gene expression during the early processes of fertilization. Also involved in regulation of receptors other than GPCRs. Involved in endocytosis of TGFBR2 and TGFBR3 and down-regulates TGF-beta signaling such as NF-kappa-B activation. Involved in endocytosis of low-density lipoprotein receptor/LDLR. Involved in endocytosis of smoothened homolog/Smo, which also requires GRK2. Involved in endocytosis of SLC9A5. Involved in endocytosis of ENG and subsequent TGF-beta-mediated ERK activation and migration of epithelial cells. Involved in Toll-like receptor and IL-1 receptor signaling through the interaction with TRAF6 which prevents TRAF6 autoubiquitination and oligomerization required for activation of NF-kappa-B and JUN. Involved in insulin resistance by acting as insulin-induced signaling scaffold for SRC, AKT1 and INSR. Involved in regulation of inhibitory signaling of natural killer cells by recruiting PTPN6 and PTPN11 to KIR2DL1. Involved in the internalization of the atypical chemokine receptor ACKR3 (By similarity). Acts as an adapter protein coupling FFAR4 receptor to specific downstream signaling pathways, as well as mediating receptor endocytosis. During the activation step of NLRP3 inflammasome, directly associates with NLRP3 leading to inhibition of pro-inflammatory cytokine release and inhibition of inflammation Functions in regulating agonist-mediated G-protein coupled receptor (GPCR) signaling by mediating both receptor desensitization and resensitization processes. During homologous desensitization, beta-arrestins bind to the GPRK-phosphorylated receptor and sterically preclude its coupling to the cognate G-protein; the binding appears to require additional receptor determinants exposed only in the active receptor conformation. The beta-arrestins target many receptors for internalization by acting as endocytic adapters (CLASPs, clathrin-associated sorting proteins) and recruiting the GPRCs to the adapter protein 2 complex 2 (AP-2) in clathrin-coated pits (CCPs). However, the extent of beta-arrestin involvement appears to vary significantly depending on the receptor, agonist and cell type. Internalized arrestin-receptor complexes traffic to intracellular endosomes, where they remain uncoupled from G-proteins. Two different modes of arrestin-mediated internalization occur. Class A receptors, like ADRB2, OPRM1, ENDRA, D1AR and ADRA1B dissociate from beta-arrestin at or near the plasma membrane and undergo rapid recycling. Class B receptors, like AVPR2, AGTR1, NTSR1, TRHR and TACR1 internalize as a complex with arrestin and traffic with it to endosomal vesicles, presumably as desensitized receptors, for extended periods of time. Receptor resensitization then requires that receptor-bound arrestin is removed so that the receptor can be dephosphorylated and returned to the plasma membrane. Mediates endocytosis of CCR7 following ligation of CCL19 but not CCL21. Involved in internalization of P2RY1, P2RY4, P2RY6 and P2RY11 and ATP-stimulated internalization of P2RY2. Involved in phosphorylation-dependent internalization of OPRD1 and subsequent recycling or degradation. Involved in ubiquitination of IGF1R. Beta-arrestins function as multivalent adapter proteins that can switch the GPCR from a G-protein signaling mode that transmits short-lived signals from the plasma membrane via small molecule second messengers and ion channels to a beta-arrestin signaling mode that transmits a distinct set of signals that are initiated as the receptor internalizes and transits the intracellular compartment. Acts as a signaling scaffold for MAPK pathways such as MAPK1/3 (ERK1/2) and MAPK10 (JNK3). ERK1/2 and JNK3 activated by the beta-arrestin scaffold are largely excluded from the nucleus and confined to cytoplasmic locations such as endocytic vesicles, also called beta-arrestin signalosomes. Acts as a signaling scaffold for the AKT1 pathway. GPCRs for which the beta-arrestin-mediated signaling relies on both ARRB1 and ARRB2 (codependent regulation) include ADRB2, F2RL1 and PTH1R. For some GPCRs the beta-arrestin-mediated signaling relies on either ARRB1 or ARRB2 and is inhibited by the other respective beta-arrestin form (reciprocal regulation). Increases ERK1/2 signaling in AGTR1- and AVPR2-mediated activation (reciprocal regulation). Involved in CCR7-mediated ERK1/2 signaling involving ligand CCL19. Is involved in type-1A angiotensin II receptor/AGTR1-mediated ERK activity. Is involved in type-1A angiotensin II receptor/AGTR1-mediated MAPK10 activity. Is involved in dopamine-stimulated AKT1 activity in the striatum by disrupting the association of AKT1 with its negative regulator PP2A. Involved in AGTR1-mediated chemotaxis. Appears to function as signaling scaffold involved in regulation of MIP-1-beta-stimulated CCR5-dependent chemotaxis. Involved in attenuation of NF-kappa-B-dependent transcription in response to GPCR or cytokine stimulation by interacting with and stabilizing CHUK. Suppresses UV-induced NF-kappa-B-dependent activation by interacting with CHUK. The function is promoted by stimulation of ADRB2 and dephosphorylation of ARRB2. Involved in IL8-mediated granule release in neutrophils (By similarity). Involved in p53/TP53-mediated apoptosis by regulating MDM2 and reducing the MDM2-mediated degradation of p53/TP53. May serve as nuclear messenger for GPCRs. Upon stimulation of OR1D2, may be involved in regulation of gene expression during the early processes of fertilization. Also involved in regulation of receptors other than GPCRs. Involved in endocytosis of TGFBR2 and TGFBR3 and down-regulates TGF-beta signaling such as NF-kappa-B activation. Involved in endocytosis of low-density lipoprotein receptor/LDLR. Involved in endocytosis of smoothened homolog/Smo, which also requires GRK2. Involved in endocytosis of SLC9A5. Involved in endocytosis of ENG and subsequent TGF-beta-mediated ERK activation and migration of epithelial cells. Involved in Toll-like receptor and IL-1 receptor signaling through the interaction with TRAF6 which prevents TRAF6 autoubiquitination and oligomerization required for activation of NF-kappa-B and JUN. Involved in insulin resistance by acting as insulin-induced signaling scaffold for SRC, AKT1 and INSR. Involved in regulation of inhibitory signaling of natural killer cells by recruiting PTPN6 and PTPN11 to KIR2DL1. Involved in the internalization of the atypical chemokine receptor ACKR3 (By similarity). Acts as an adapter protein coupling FFAR4 receptor to specific downstream signaling pathways, as well as mediating receptor endocytosis. During the activation step of NLRP3 inflammasome, directly associates with NLRP3 leading to inhibition of pro-inflammatory cytokine release and inhibition of inflammation |
|---|---|
| 基因 ID | 409 |
| 基因名 | ARRB2, Arrb2 |
| Swiss | P32121, Q91YI4, P29067 |
| 别名 | Beta Arrestin 2 (YD16141),Beta Arrestin 2 (YD16141) Rabbit mAb,ARRB2,Arrestin beta-2,Non-visual arrestin-3,ARB2,ARR2 |
资料与技术支持
验证数据
1 张验证图
页面顶部轮播可查看应用验证图,覆盖 WB / FC / IP。
常见问题
当前页面标注应用包括 WB, FC, IP,建议结合页面验证图和推荐稀释比例进行预实验优化。
可通过页面询价/留言入口提交货号和批号,技术支持会协助提供对应批次资料。
页面推荐条件可作为起始浓度,不同样本、固定方式和检测体系可能需要梯度优化。