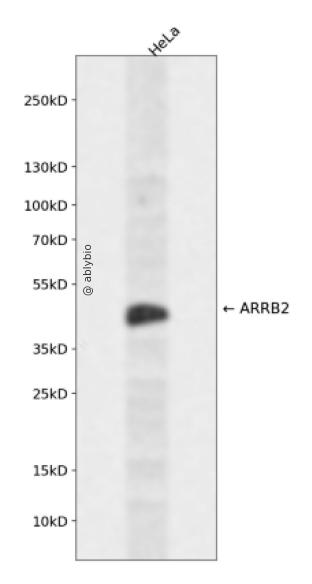
Western blot analysis of ARRB2 expressed in HeLa using ARRB2 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
Western blot analysis of ARRB2 expressed in HeLa using ARRB2 Rabbit pAb at 1:1000. Secondary antibody: HRP Goat Anti-Rabbit IgG (H+L) at 1:5000. Lysates/proteins: 30ug per lane. Blocking buffer: 5% non-fat dry milk in TBST. Detection: ECL Enhanced Kit. Exposure time: 120s.
Members of arrestin/beta-arrestin protein family are thought to participate in agonist-mediated desensitization of G-protein-coupled receptors and cause specific dampening of cellular responses to stimuli such as hormones, neurotransmitters, or sensory signals. Arrestin beta 2, like arrestin beta 1, was shown to inhibit beta-adrenergic receptor function in vitro. It is expressed at high levels in the central nervous system and may play a role in the regulation of synaptic receptors. Besides the brain, a cDNA for arrestin beta 2 was isolated from thyroid gland, and thus it may also be involved in hormone-specific desensitization of TSH receptors. Multiple alternatively spliced transcript variants encoding different isoforms have been found for this gene.
Functions in regulating agonist-mediated G-protein coupled receptor (GPCR) signaling by mediating both receptor desensitization and resensitization processes. During homologous desensitization, beta-arrestins bind to the GPRK-phosphorylated receptor and sterically preclude its coupling to the cognate G-protein; the binding appears to require additional receptor determinants exposed only in the active receptor conformation. The beta-arrestins target many receptors for internalization by acting as endocytic adapters (CLASPs, clathrin-associated sorting proteins) and recruiting the GPRCs to the adapter protein 2 complex 2 (AP-2) in clathrin-coated pits (CCPs). However, the extent of beta-arrestin involvement appears to vary significantly depending on the receptor, agonist and cell type. Internalized arrestin-receptor complexes traffic to intracellular endosomes, where they remain uncoupled from G-proteins. Two different modes of arrestin-mediated internalization occur. Class A receptors, like ADRB2, OPRM1, ENDRA, D1AR and ADRA1B dissociate from beta-arrestin at or near the plasma membrane and undergo rapid recycling. Class B receptors, like AVPR2, AGTR1, NTSR1, TRHR and TACR1 internalize as a complex with arrestin and traffic with it to endosomal vesicles, presumably as desensitized receptors, for extended periods of time. Receptor resensitization then requires that receptor-bound arrestin is removed so that the receptor can be dephosphorylated and returned to the plasma membrane. Mediates endocytosis of CCR7 following ligation of CCL19 but not CCL21. Involved in internalization of P2RY1, P2RY4, P2RY6 and P2RY11 and ATP-stimulated internalization of P2RY2. Involved in phosphorylation-dependent internalization of OPRD1 and subsequent recycling or degradation. Involved in ubiquitination of IGF1R. Beta-arrestins function as multivalent adapter proteins that can switch the GPCR from a G-protein signaling mode that transmits short-lived signals from the plasma membrane via small molecule second messengers and ion channels to a beta-arrestin signaling mode that transmits a distinct set of signals that are initiated as the receptor internalizes and transits the intracellular compartment. Acts as signaling scaffold for MAPK pathways such as MAPK1/3 (ERK1/2) and MAPK10 (JNK3). ERK1/2 and JNK3 activated by the beta-arrestin scaffold are largely excluded from the nucleus and confined to cytoplasmic locations such as endocytic vesicles, also called beta-arrestin signalosomes. Acts as signaling scaffold for the AKT1 pathway. GPCRs for which the beta-arrestin-mediated signaling relies on both ARRB1 and ARRB2 (codependent regulation) include ADRB2, F2RL1 and PTH1R. For some GPCRs the beta-arrestin-mediated signaling relies on either ARRB1 or ARRB2 and is inhibited by the other respective beta-arrestin form (reciprocal regulation). Increases ERK1/2 signaling in AGTR1- and AVPR2-mediated activation (reciprocal regulation). Involved in CCR7-mediated ERK1/2 signaling involving ligand CCL19. Is involved in type-1A angiotensin II receptor/AGTR1-mediated ERK activity. Is involved in type-1A angiotensin II receptor/AGTR1-mediated MAPK10 activity. Is involved in dopamine-stimulated AKT1 activity in the striatum by disrupting the association of AKT1 with its negative regulator PP2A. Involved in AGTR1-mediated chemotaxis. Appears to function as signaling scaffold involved in regulation of MIP-1-beta-stimulated CCR5-dependent chemotaxis. Involved in attenuation of NF-kappa-B-dependent transcription in response to GPCR or cytokine stimulation by interacting with and stabilizing CHUK. Suppresses UV-induced NF-kappa-B-dependent activation by interacting with CHUK. The function is promoted by stimulation of ADRB2 and dephosphorylation of ARRB2. Involved in p53/TP53-mediated apoptosis by regulating MDM2 and reducing the MDM2-mediated degradation of p53/TP53. May serve as nuclear messenger for GPCRs. Upon stimulation of OR1D2, may be involved in regulation of gene expression during the early processes of fertilization. Also involved in regulation of receptors other than GPCRs. Involved in endocytosis of TGFBR2 and TGFBR3 and down-regulates TGF-beta signaling such as NF-kappa-B activation. Involved in endocytosis of low-density lipoprotein receptor/LDLR. Involved in endocytosis of smoothened homolog/Smo, which also requires GRK2. Involved in endocytosis of SLC9A5. Involved in endocytosis of ENG and subsequent TGF-beta-mediated ERK activation and migration of epithelial cells. Involved in Toll-like receptor and IL-1 receptor signaling through the interaction with TRAF6 which prevents TRAF6 autoubiquitination and oligomerization required for activation of NF-kappa-B and JUN. Involved in insulin resistance by acting as insulin-induced signaling scaffold for SRC, AKT1 and INSR. Involved in regulation of inhibitory signaling of natural killer cells by recruiting PTPN6 and PTPN11 to KIR2DL1. Involved in IL8-mediated granule release in neutrophils. Involved in the internalization of the atypical chemokine receptor ACKR3.
a. 贴壁培养细胞收集
去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
b. 悬浮培养细胞收集
速离心悬浮细胞,弃上清,收集沉淀。手指轻弹细胞,使其松散。
c. 组织样本收集
把组织剪切成细小的碎片,越小越好。取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
(2)总蛋白提取
a. 细胞/组织裂解
将装有细胞沉淀或组织碎片的容器完全插入冰中。细胞沉淀按照1mL裂解液/107个细胞(1个T75培养瓶细胞量)的比例加入相应体积的裂解液(细胞量足够时都加入3mL,不足时根据细胞量计算),裂解20min,每隔5min将离心管置于涡旋振荡仪上震荡10s。组织碎片按照0.5mL 裂解液/100mg组织向匀浆器中加入蛋白裂解液,每3min研磨一次,重复5次,使组织尽量碾碎。(裂解液中根据需要选择添加或不添加蛋白酶抑制剂)。
b. 离心
把裂解好的样品配平后,置于预冷的高速冷冻离心机中,12000 rpm,15min。
c. 蛋白变性
完成离心后,上清即为蛋白提取液。吸取少量蛋白提取液做蛋白浓度测定。向剩余的蛋白提取液的离心管中加入1/5上清体积的5×Loading Buffer(最终工作液为1X),待干式恒温器温度升至95℃后,将1.5mL离心管插入加热孔中,95℃加热变性10min,待液体完全冷却后置于-20℃保存。
(3)蛋白浓度测定(BCA法)
a. BCA工作液的配置
根据样品数量,按50体积BCA试剂A加入1体积BCA试剂B(50:1)配置适量BCA工作液,充分混匀。BCA工作液室温24h内稳定。
b. 标准品测定
取10μl蛋白标准品(5mg/ml BSA)稀释至50μl,使终浓度为1mg/ml。稀释后的蛋白标准品可以-20℃长期保存。此标准品溶液的稀释液可使用去离子水或1*PBS。将标准品按0、1、2、4、8、12、16、20μl加入到96孔板中,加稀释液补足到20μl(见附表)。加适当体积样品到96孔板的样品孔中,如果样本不足20μl,需加稀释液补足到20μl。请注意记录样品体积。各孔加入200μl BCA工作液,37℃放置20-30min。用酶标仪测定A562,或540-595nm之间的其他波长吸光度。根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
a. Western Transfer Buffer至少提前2h (即开始电泳后)放入-20℃冰箱预冷,但注意避免结冰。
b. 根据胶体大小,将Filter Paper及Nitrocellulose membrane剪裁至合适尺寸。
c. 目的蛋白>20KD选择0.45μm NC膜/PVDF膜;目的蛋白<20KD选择0.2μmNC膜或PVDF膜,选择完毕后将NC膜放在Western Transfer Buffer中浸泡备用,注意如使用的是PVDF膜需先放入甲醇中浸泡5-10min,再放入Western Transfer Buffer中浸泡备用。
(2)裂解液&洗杂液:Cell lysis buffer for IP (without inhibitors)
(3)蛋白酶抑制剂
(4)封闭液:含 3% BSA 的 1X PBS
(5)1×PBS 缓冲液
(6)5×loding buffer(使用时用去离子水稀释至工作浓度即可)
(7)Control IgG (AC005/ AC011/AC034)
二、实验步骤
1、样本处理
(1)贴壁培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 去除贴壁细胞的培养液,用PBS、NS或无血清培养基清洗1次,低速离心,弃上清,留取沉淀。
c. 按照6孔板每孔加入100~200μl裂解液的比例,加入裂解液。移液器轻轻吹打,使裂解液和细胞充分接触。通常裂解液作用于细胞1~5s内,细胞会被裂解。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(2)悬浮培养细胞
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 速离心悬浮细胞,弃上清,收集沉淀。
c. 手指轻弹细胞,使其松散。按照6孔板每孔加入100~200μl裂解液的比例,加入NP-40裂解液。通常6孔板每孔加入100~200μl裂解液已经足够,但如果细胞密度非常高可以适当加大裂解液的用量150~200μl,再用手指轻弹以充分裂解细胞。充分裂解后应无明显沉淀。
d. 1000~12000g,离心3~5min(如果用冷冻离心机4℃效果更佳),取上清。
(3)组织样本
a. 取裂解液室温溶解混匀,根据需要选择添加或不添加蛋白酶抑制剂。
b. 把组织剪切成细小的碎片,越小越好。
c. 取液氮或超低温冰箱中冷冻30min以上的组织,迅速用液氮研磨,研磨过程尽量控制在1~2min之内,以减少蛋白的降解。
d. 按照每20mg组织加入100~200μl裂解液的比例,加入含有PMSF的裂解液。冰上或4℃裂解30-60min。(步骤3、4也可采用以下过程:按照每20mg组织加入100~200μl裂解液的比例加入NP-40裂解液。用玻璃匀浆器或组织研磨器匀浆,直至充分裂解,过程尽量控制在1~2min之内,以减少蛋白的降解。)
e. 按照每20mg组织加入100~200μl裂解液的比例,加入裂解液。
f. 1000~12000g,4℃离心10~15min(如无低温离心机,室温下离心也可),取上清。
2、磁珠预处理
(1)将rProtein A/G Plus MaqPoly Beads颠倒或漩涡混匀,翻转瓶身发现底部无黑色沉淀即可。
(2)取30μl rProtein A/G Plus MaqPoly Beads至新的EP管中,放在磁分离器上,待溶液澄清后,用移液器吸弃保护液。
(3)将EP管从磁分离器上取下来,加入1ml Cell lysis buffer for IP (without inhibitors),混匀,放置在磁分离器上,收集磁珠,用移液器吸弃洗杂液,重复2次。